

Urbanization is arguably one of the most rapid and conspicuous human-driven environmental changes, and fragmentation is one of the most pervasive outcomes of urbanization. Patches of vegetation within the built matrix of a typical cityscape, such as parks and gardens, are usually small and isolated and movement among them potentially jeopardized. Bumblebees are common in cities, which they seem to prefer to the even more unnatural modern agricultural landscape. Most of them are social and they are important pollinators of many wild plants and commercial crops. They show a great intraspecific variation in body size, a trait known to enhance the ability of organisms to move across long distances, an advantage in fragmented environments. This led us to ask whether bumblebees are actually larger in cities, something we expected on the account that bigger bees can fly the greater distances that the fragmented cityscape demands. Larger bumblebees are also better pollinators, as they are capable of depositing more pollen grains on flower stigmas. We therefore also asked whether urban fragmentation, by virtue of its potential effect on body size, might also have an indirect effect on pollination.
Study Species, Sampling Locations & Methods
For the purpose of our study, we used three widespread bumblebee species, the red-tailed bumblebee Bombus lapidarius and the common carder bee B. pascuorum, which are known to forage close to their nests, and the long distance forager, the buff-tailed bumblebee B. terrestris. We sampled more than 1,800 bumblebee workers from 18 sites located in nine German cities and in nine paired rural locations (Figure 1). We sampled exclusively in flower-rich sites in both the rural and urban ecosystem, to avoid the confounding effect of local floral resource availability on body size. We then measured the intertegular distance (ITD, proxy for body size) of every bumblebee we collected. We additionally calculated the percentage coefficient of variation of ITD and the community-weighted mean (CWM) and community-weighted mean coefficient of variation (CWM CV) of bumblebee body size.
To determine the main environmental correlates of body size, we quantified flowering plant abundance, temperature and wind speed. Using land cover maps, we also quantified road density at multiple spatial scales (250, 500, 750 and 1000 m) as a metric of urban fragmentation.
At each sampling site we also evaluated the pollination success of red clover, Trifolium pratense, a self-incompatible plant species that is preferentially visited by bumblebees. Red clover plants were grown in an insect-free glasshouse (Figure 2). Potted plants, each with the same number of marked open inflorescences, were then placed at each field site and then returned to the insect-free greenhouse until seeds were formed. Seeds were counted and the average number of seeds per plant was used as a measure of the ecosystem service of pollination.

Urbanization had a positive effect on bumblebee body size
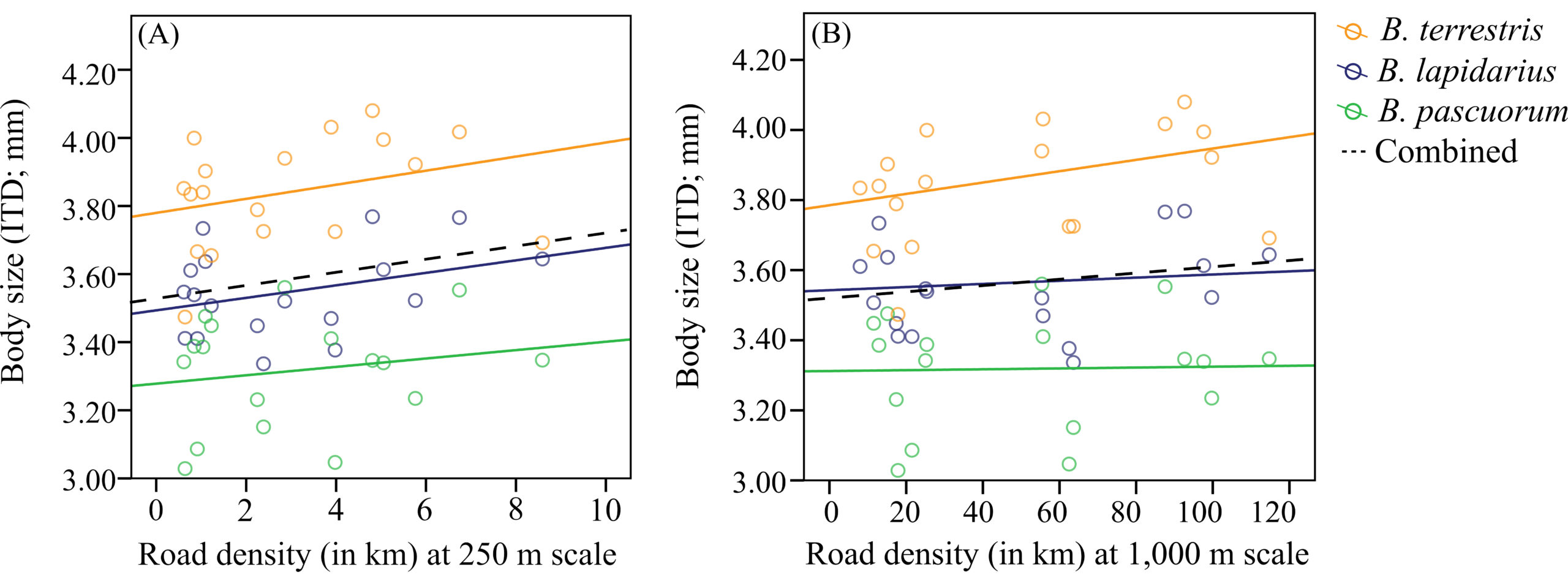
The mean body size of all bumblebee species increased with increasing road density however, at different scales. The body size of the bumblebees with shorter foraging ranges (Bombus lapidarius and B. pascuorum), was positively correlated with road density at 250 m, while the body size of the long distance forager, B. terrestris, was positively correlated with road density at all scales (Figure 3). These results have two important implications: 1) given that road density is higher in urban sites, it confirms urbanisation-mediated fragmentation as the factor to which these bees respond across the urban/rural divide; 2) the scale at which our three bumblebee study species perceive the landscape and respond to environmental challenges is different, probably because of their different absolute body size (Bombus terrestris being larger than the two other species).
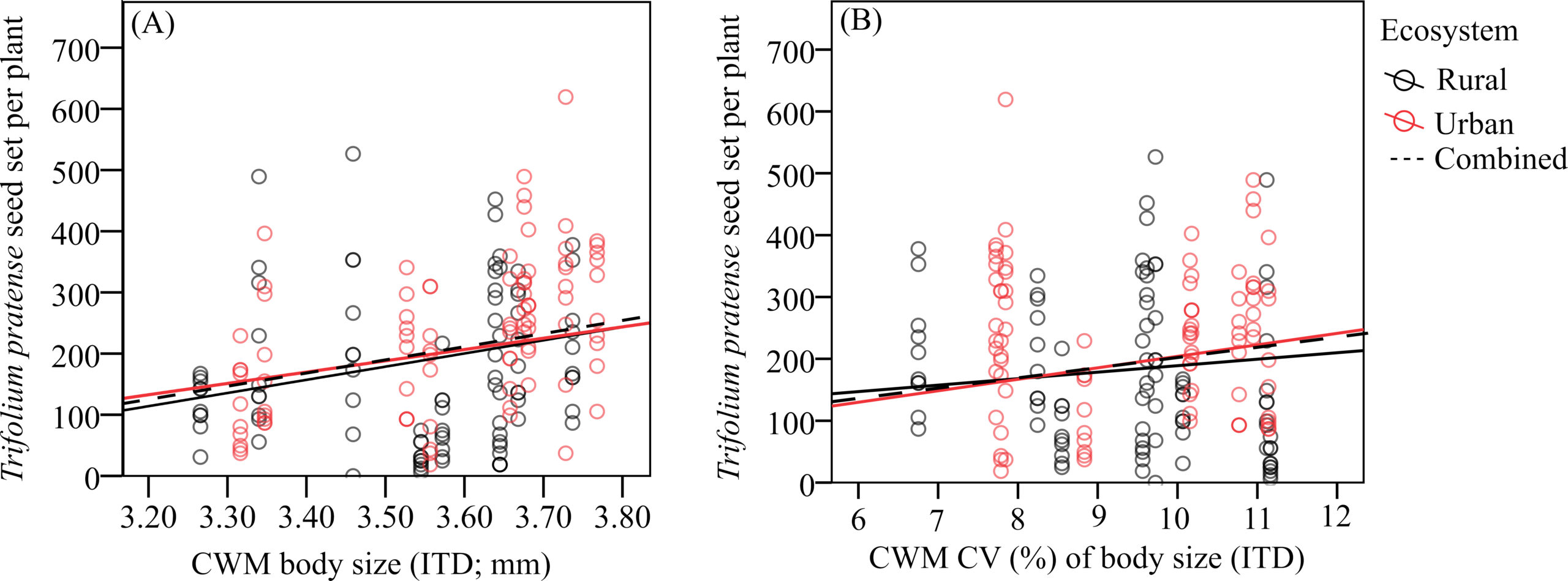
Urbanization had an indirect, positive effect on pollination through its effects in boosting bumblebee community mean body size and its variation
Urbanization had a positive effect on pollination service provision via its direct effects on bumblebee body size distribution. We confirmed body size as a predictor of pollination effectiveness (Figure 4) and explicitly link urbanisation to a trait (body size) that affects a key ecosystem function (pollination), with potential repercussions at the ecosystem level. These results are even more important given that bumblebees are the dominant pollinators, which makes the understanding of their potential to respond to environmental change even more crucial.
Eco-evolutionary implications
In bees, body size is a highly plastic trait, with low heritability and major effects on fitness. Within a bumblebee colony, workers, despite being highly related, display as much as a 10-fold difference in body size. We detected a body size distribution shift between rural and urban ecosystems, which was due to bumblebees being overall larger in cities. This environmentally-driven morphological change might have arisen as a result of selection and/or phenotypic plasticity. But for mobile organisms such as bumblebees, which have the opportunity to move through the landscape to find the environmental conditions that best match their phenotype, habitat matching might also be involved. We cannot confirm or exclude any of these processes. We argue, that the colony level bumblebee variability in body size confers a colony the plasticity needed to adjust to the fragmentation imposed by urban settings. The question around the importance of plasticity in evolution is not new and debate continues as to whether plasticity facilitates or hampers evolutionary change. Equally for bumblebees, the question of whether the body size shift we observed has already or will translate into evolutionary change (that is, a change in allele frequency) is an open question that can be addressed only through carefully designed experiments and concomitant genetic analyses.
Featured Image: “Bumblebee (Bombus lapidarius), Sandy, Bedfordshire” by orangeaurochs is licensed under CC BY 2.0
This post was co-authored by: [starbox id=68,69]
Leave a Reply