
Pathogens are organisms that cause disease, and zoonotic pathogens are those that can be transmitted between animals and people. Where pathogens are found can be determined by environmental changes. Urbanization is known to mediate environmental changes in cities as well as impact biodiversity. In Brazil, Belo et al. (2011) found a higher prevalence of parasites from the genera Plasmodium and Haemoproteus in an urban site when compared to non-urban areas. This pattern varies across taxa and can even differ within taxa. Therefore, understanding how urban areas influence pathogen distribution is crucial for management of health risks across cities. For example, the density of structures within urban systems can affect the spatiotemporal distribution of individuals which translates to the density of particular pathogens. This can influence what pathogens are successful within specific taxa that are successful in urban environments.
How do pathogens currently exist in cities?
Rats serve as a source for a number of pathogens. These pathogens can be spread directly or indirectly in several ways (e.g., fleas, rat bites, and urine). Given the large distribution of rats and their ability to successfully adapt to cities, understanding rat-pathogen dynamics is crucial, especially in areas of intense urbanization.
Previous studies on Norway rats showed that relatedness between individuals varies over space and that genetic methods can reveal the movements of individuals. This gives researchers insight into how pathogens spread or cluster in cityscapes. Recently, Byers et al. set out to understand more about the distribution of three pathogens of health concern (Leptospira interrogans, Bartonella tribocorum, and Clostridium difficile), combining previously published disease and genomic data to investigate the distribution of pathogens in rats and how these patterns compare to genetic structure. These researchers were interested in a) identifying closely related individuals to infer movement, b) comparing patterns of relatedness and movement to prevalence data for pathogenic bacteria carried by rats, and c) exploring the impact of family members on an individuals’ pathogen status.
Methods
Rats were trapped and humanely euthanized to collect the kidney, blood, and colon contents for L. interrogans, B. tribocorum, and C. difficile, respectively. To reveal the relatedness between individuals, rats were genotyped and related rats were identified using the R package Sequoia which identifies relatives such as parents and their offspring, as well as siblings. To evaluate whether genetic relatedness influences pathogen status, a binomial generalized linear mixed model was used. To infer travel distance of individuals, the Euclidian distance between a set of relatives was calculated and categorized by relationship type.
Results and Conclusion
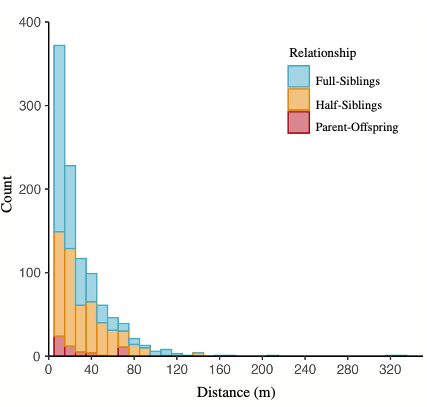
Using rat genetics, Byers et al. identified 1,246 pairs of relatives and found that these related pairs were caught from 0-330 m apart. Of these pairs, 75% of related individuals were caught within 33 m, with full-siblings being caught closest to each other (median = 10 m) and half-siblings being the furthest apart (median = 22.5 m) (Figure 3). This study revealed the spatial clustering of related individuals within 33m and 99% of relatives were caught within the same city block (Figure 2). This distinct clustering reflects previous studies showing that urban rats are territorial with home ranges of 30 – 45 m in diameter. And although rats can travel long distances, this is rare. This, along with the clustering, suggest very little movement between city blocks, resulting in low interactions between rats in neighboring blocks and little opportunity for pathogen spread.
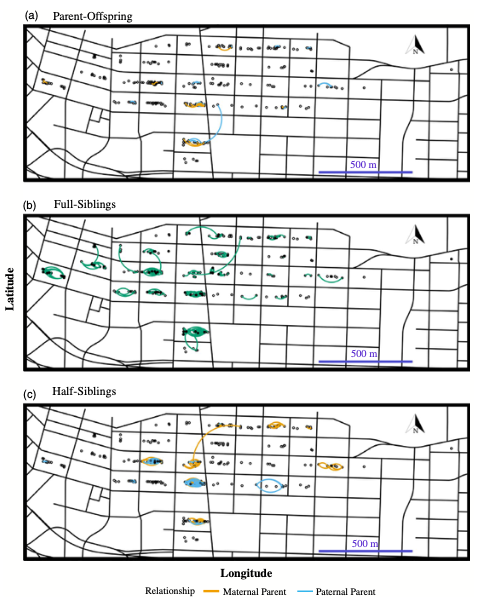
The researchers also find that pairs of blocks with relatives often share the same pathogen status (e.g.., affected or not). However, having a full-sibling with a pathogen did not seem to influence a rat’s carriage of B. tribocorum or L. interrogans. These pathogens were expected to be found within a full-sibling family because of how they are transmitted (i.e., fleas and urine) which requires near contact between individuals.
Overall, their work demonstrates the block-level association may be more important to consider when evaluating patterns of prevalent pathogens than full-sibling relatedness. This work demonstrated that related rats often aggregate within 33 m, which is particularly relevant for the management of outbreaks via rat-associated health-risks.
Read the full article here.
Literature referenced:
Belo NO, Pinheiro RT, Reis ES et al (2011) Prevalence and lineage diversity of avian haemospo- ridians from three distinct cerrado habitats in Brazil. PLoS One 6:e17654
This post was co-authored by: [starbox id=67,65]
Leave a Reply